Thistles
Thistles are produced in a Thistle Garden, and they are used to feed Sheep and Silkworms. Raw Silk, from the silkworms, is subsequently used to make Silk Cloth which is a valuable luxury item.- Modern Sheep Farms use both Onions and Thistles as feed stock. For thistles, two vitamins will be required (~ or +), and the other five vitamins can have any level. You must provide at least one thistle with that specification, as that is the only way (other than rebuilding) to change the requirements for the next batch.
- Silkworm Farms hatch new silkworms each game week. Once the silkworms are born, an avatar with the Silkworm Farming skill (level 28) can determine the required level of four different vitamins that the current batch of silkworms will need in order to grow. The other three vitamins can have any level.
 |  |  |
| Thistle Gardens | Modern Sheep Farm | Silkworm Farm |
Vitamins
Seven vitamins are defined:- A - Asc - Ascorbin
- B - Bio - Biotin
- C - Car - Carotine
- F - Fol - Folin
- N - Nia - Niacin
- P - Pyr - Pyroxiding
- T - Thi - Thiamine
Vitamins can have concentrations that range from 0-99, but the requirements are specified as A+B-C~N+, for example, without indicating the unrestrained vitamins. The vitamin levels shown on the Thistle Garden control widget appear to be real numbers that are truncated, not rounded, and those numerical vitamin levels are further evaluated and inventoried as only one of three states for each vitamin, - < 21, ~, 79 < +.
There are 3 possible states for each of 7 different vitamins, and therefore 3*7=21 distinct states for any particular thistle if we ignore combinations. For Modern Sheep Farms the required thistle must have ~ or + values for two specified vitamins. If we pretend that all of the vitamin states are equally probable, and it's actually pretty easy to force a couple of vitamins above 21, the odds that a thistle which has the first vitamin also has the second vitamin is 4/21 * 2/18 = 2%. If you have 50 varieties of thistle you'll probably have what you need to feed the sheep. Sheep are easy to feed.
Silkworms are more difficult. Four of the vitamins must have a particular state. As each of the seven vitamins can be in one of three different states, the number of possible thistle specifications for feeding the silkworms is somewhat less than 3^7=2187, because -~+ are exclusive states and only 4 of the 7 Vitamins are restrained. Your odds that a particular thistle will have everything that you need should be 4/21 * 3/18 * 2/15 * 1/12 = 0.035%. If you had a random assortment of thistles you'd need 2835 different varieties on hand, so it's not practical to warehouse thistles nor to list every possible recipe, especially so when environmental conditions may allow various recipes to reach a given target state. We will therefore emphasize:
- How to reliably determine the available conversions
- How to traverse the consequent state diagram to obtain the required vitamin levels
Nutrients
Five nutrients are defined:- D - Nit - Nitrogen, from (Sheep or Camel) Dung
- U - Pot - Potassium, from Saltpeter, from (Sheep or Camel) Urine (even though the game calls it Dung)
- H - H2O - Water, applied directly, using a clay Jug full of water
- O - Oxy - Oxygen, from aerating the soil (No cultivator nor any other special tool is needed.)
- S - Sun - Sunlight, determined by time-of-day and the position of the canopy
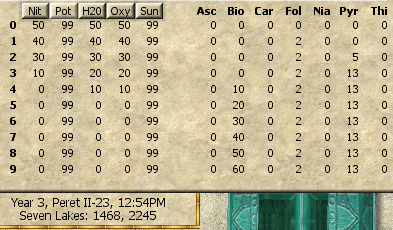
- You plant a thistle seed to start the process. All of the ingredients except Sunshine initialize at 50%, so you get some free Dung (Nitrogen), Urine (Potassium from Saltpeter), H2O (Water), and Oxygen.
- While free may seem like a good deal, in reality the free ingredients just mess you up by introducing a bunch of undesired variables. Growing thistles would be much simpler if there were no free ingredients. Even if you blow off the first 5 steps to zero out the ingredients, you'll usually have some stray vitamins that you have to accommodate.
- The amount of available Sunlight varies by time-of-day, with a maximum value of 99% for six hours during mid-day, gradually diminishing to a minimum value of zero from 6:40pm until 5:20am the following morning, and then increasing again to the maximum value.
- Closing (toggling) the canopy blocks two/thirds of the available Sunlight.
- Thistle growth occurs in 40 steps of 6 seconds each.
- During each step you can add Dung (Nitrogen), Urine (Potassium from Saltpeter), H2O (Water, so you'll need some clay Jugs), or Oxygen (from aeration of the soil, but you don't need a cultivator or any special tools) in increments of 20 units, to a maximum value of 99 units.
- Sunshine is more complicated, as the amount of available sunshine depends on the time-of-day and on whether or not the canopy is Open (all available sunshine) or Closed (1/3 of the available sunshine).
- During each step the level of Dung or Urine will decrement by 10, 20, or 30 units, depending on the type and the number of active conversions. The level of H2O and Oxygen will always decrement by -10 units. Sunlight only depends on the time-of-day and the position of the canopy.
- Ingredients are added 20 units at a time, so it's not possible to hold them at a specific intermediate value.
- If you add material when the value is already at 79 units or more, the value will cap at 99 units
- You can gradually increment (-10+20)= +10 units per step by adding 20 units of material during each step
- You can gradually decrement -10 units per step by doing nothing during each step
- Dung, Urine, H2O, and Oxygen can only take 20 integer values (0, 9, 10, 19, 20, ..., 79, 80, 89, 90, 99).
- Only even numbers are available until you max out the value
- Only odd numbers are available thereafter
- Sunshine can have any integer value (it's actually a truncated real value) from 0 through 99.
- Because the conversion rates are fractional, Vitamins can show any integer value from 0 through 99.
- The amount Dung and Urine on hand doesn't seem to matter, so long as you have enough in the hopper to support the currently active conversions.
- The state calculations seem to be local. In other words, how you got to a particular state doesn't matter. What you have right now, and the available conversions, completely defines the next set of values.
- Regardless of which buttons you push (other than aborting), on the 40th step the conversions will cease and you can then harvest 2 seeds plus 5 thistles of some sort.
Sunshine
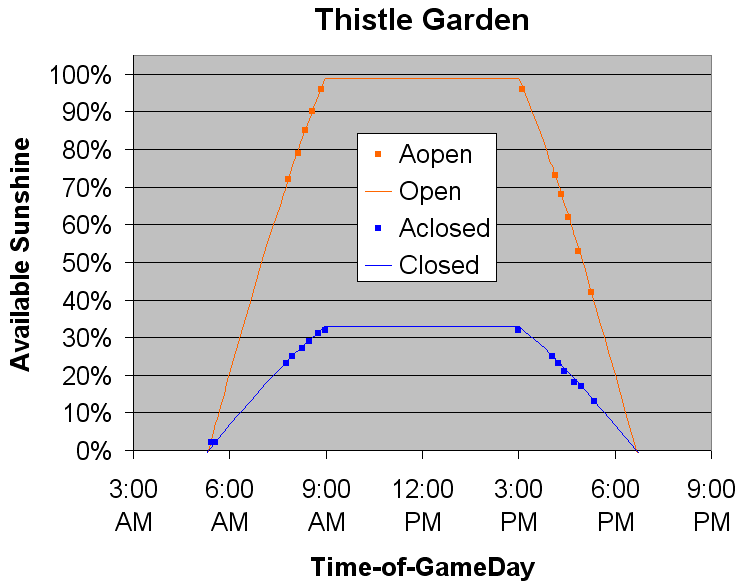
As you can well imagine, the amount of sunlight is an important factor that influences which conversions are available, and also the rate of conversion in some cases. Controlling the amount of Sunlight isn't tricky, you just have to grow thistles at the appropriate time of day for what you want to accomplish.
The available sunshine is a tinkered sinusoidal, with an amplitude of 1.3, a cropped period of 26.6 hours, and some clamping. The available sunshine when the canopy is closed is always 1/3 of the available sunshine when the canopy is open.
- Mid-day clamps to 99% for six hours (or 33% if the canopy is closed)
- 3:00 PM to 6:40 PM gradually decreases to zero
- Night clamps to zero for 10 hours and 40 minutes
- 5:20 AM to 9:00 AM gradually increases to 99% (or 33% of that value if the canopy is closed)
It doesn't seem to matter exactly how you managed to catch the right amount of available sunshine. Opening the canopy at 6:00am or 6:00pm is the same 20% as closing the canopy at 7:20am or 4:40pm.
- 99 for six hours during mid-day with the canopy open
- 98 to 34, each for 2.2 minutes (19.8 iterations) during morning/afternoon open
- 33 for six hours during mid-day closed or for 2.2 minutes during morning/afternoon open
- 32 to 1, each for 2.2 minutes during morning/afternoon closed or for a slightly earlier/later 2.2 minutes during dawn/dusk open
- 0 for 10+ hours during mid-night darkness, regardless of whether the canopy is open or closed
Conversions
We'll be using an analytical technique known as a Hidden Markov Model. It's an essential method for discerning the Thistle conversions, as many of them are not directly visible.
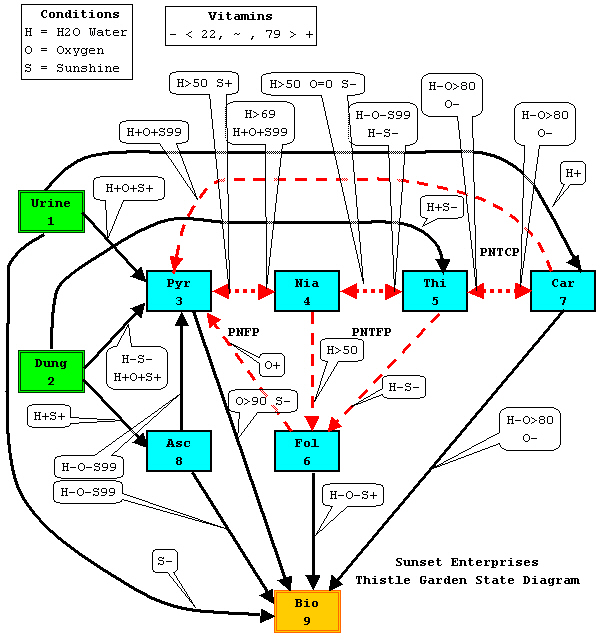
WARNING: These conversions were applicable during Tale 1 Thistles. The conversions have definitely changed for Tale 3; how much they've changed is a part of what we're testing.
The conversion relationships are what makes Thistle Cultivation an interesting puzzle. Some observations:
- Dung (Nitrogen) and Urine (Saltpeter for Potassium), on the left of the diagram, are pure inputs.
- Bio is a sink. There is no way to remove Bio.
- The conversions between Pyr <-> Nia <-> Thi <-> Car (shown as dotted red lines) are all reversible.
- Nia -> Fol and subsequently Fol -> Pyr (shown as dashed red lines) permits a state loop.
- Thi -> Fol and subsequently Fol -> Pyr (shown as dashed red lines) permits a state loop.
- Car -> Pyr (shown as a dashed red line) permits a state loop.
- The call-out balloons indicate the environmental conditions that are required for a conversion to function.
There are 20 known conversions, with some multiples for the same pairs, but only 80% of them are available on any given game day. The weather changes at midnight.
- Dung and Urine are true inputs. If you have neither in the hopper, the related conversion won't happen.
- H2O, Oxy, and Sun are not inputs, per se, rather they are environmental variables that effect all conversions concurrently.
- Vitamins can source other vitamins, and there are some equilibrium equations (the same amount is arriving and leaving) that can mask the participation of one or more vitamins.
- Some conversions proceed nicely without either Dung or Urine.
- The 50% free initial levels will leave some junk vitamins that will usually mess up your analysis, but which can occasionally be used to bootstrap a recipe.
The puzzle then, is to solve a maze with several interesting wrinkles.
- Each day some of the conversions (links) simply won't work, no matter what you do.
- Each conversion, if it is working, will only work if you have the right combination of H20, Oxy, and Sun.
- Ingredients are converted at various ratios, ranging from 3:1 through 1:5.
- Some of the links are reversible, and you'll get equilibrium conditions.
- Some of the links form loops, which can be used to advantage.
- You can steer the conversions toward your goal by adjusting the levels of Urine, Dung, H2O, Oxy, and Sun.
- Vitamins accumulate at the nodes.
The solution is not to traverse the maze, but rather to obtain the desired quantity of vitamins at four different nodes.
Research Objectives
Determine (verify) all possible conversions
Thistle cultivation hasn't really been examined since Tale 1, and the current science if pretty iffy.
We're using Hypersnap to record and print the data from each test.
The data tables indicate the Game Date (different days can provide different results), followed by Step, Ingredients, and Vitamins arranged exactly as they are presented on the screen.
The sort order for the significant states is in descending binary order from 11111 to 00000, with 1 for any value > 0 and 0 for 0. Dung and Urine are treated as available or not available, regardless of their actual values, but within each binary category of tests the other ingredients are further sorted in descending order by their actual values.
The descending sort order is
- Category (binary DUHOS)
- H20
- Oxygen
- Sunshine
- Game Date
- Iteration
It's very important to recognize which vitamins are dynamic, and which vitamins are static. If you take a look at the associated spreadsheet, we're tracking the difference between the indicated state and the subsequent state. In other words, the Thistles.xls spreadsheet allows us to discern how each state should behave, regardless of how we got to that state, by using autofiltering to select states that may be due to the same conversion.
We'll use the convention of showing -nn (a negative value) to indicate that a particular value has stabilized, while nn (a positive value) indicates that the value will be different on the next step. Although the vitamin values are never actually negative, the convenient use of a negative for static values allows us to use basic auto-filtering tools to exclude the variables that are not effected by the current conversions (or are perhaps in equilibrium).
Initialization Tests
Our first set of tests will treat Dung, Urine, H2O, and Oxygen as binary values. We'll either maximize a value (99) or let it decrement to its minimum value (0). Those values will be clamped until the conversion rates stabilize, whereupon we'll snapshot the test results and leave the thistle garden to complete it's cycle.As each test starts with all ingredients at 50%, five steps are sufficient to zero out any undesired ingredients (but you'll very likely still have some stray vitamins). We'll therefore snapshot the initialization tests at step 6 and then move on to other tests rather than waiting for the predictable conversions to reach a maximum value.
Part of the initialization test regime will be to determine just how sensitive the various conversions are to Available Sunshine, and to define equivalent ranges. Take a close look at the test series for 0,99,99,99,S. We'll initially track Available Sunshine in the following ranges:
- Full Sunshine = 99
- High Sunshine 33 > + < 99 (This could be ugly. We may need a Medium Sunshine range as well.).
- Shaded Sunshine = 33
- Low Sunshine 0 > - < 33 and the level will be considered as equivalent whether due to time-of-day or canopy condition. (This could also be ugly.)
- Darkness with Sunshine = 0
To reiterate, for this set of initialization tests Dung, Urine, H2O, and Oxygen will be either minimized or maximized, while Sunshine will be categorized as 0%, 1%-32%, 33%, 34%-98%, or 99%. We'll only worry about five different sunlight conditions unless test results suggest that other breakpoints are significant.
The initialization test set therefore has 2*2*2*2*2=32 + 2*2*2*2*3=48 = 80 differentiated states. If multiple values are shown for a particular state it's because the weather changed, and the date with the highest number of active conversions will be bolded.
Do please note that only the tests for Iteration 6 are initialization tests. Initialization tests should be entirely reproducible. At iteration 0, immediately clamp any ingredient value that shows as 99 and keep it at 99, while letting the rest of of the values drift down to 0. If you get different tests results, it's because different conversions were available on that day.
Where an iteration other than 6 is indicated, the ingredient data serves primarily as an index to the one or more vitamin conversions are of interest. It is likely that getting to the indicated state required multiple strategies or intermediate ingredient values, and without going back to the original recipe (Thank God for Hypersnap!) it's not likely that you would accidently get the exact same results.
If you take a look at the attached Thistles.xls spreadsheet, we're also tracking the rate of change for each of the vitamins. How much of each vitamin has accumulated is nice to know, and it's actually the ultimate objective, but for research purposes the critical datum is the Rate of Change, and thereby the discernment of the relevant conversion pairs and conditions.
| GameDate | I | Nit | Pot | H2O | Oxy | Sun | Asc | Bio | Car | Fol | Nia | Pry | Thi | Ambient Conditions For This Conversion |
| 3 Peret III-19 | 6 | 99 | 99 | 99 | 99 | 33 | 19 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret II-26 | 6 | 99 | 99 | 99 | 99 | 5 | 0 | 15 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret III-28 | 6 | 99 | 99 | 99 | 99 | 0 | 0 | 15 | 19 | 0 | 39 | 0 | 0 | Car U O99 Nia H99 |
| 3 Peret III-19 | 6 | 99 | 99 | 99 | 0 | 99 | 0 | 0 | 0 | 0 | 8 | -3 | 0 | Nia H99 |
| 3 Peret III-19 | 6 | 99 | 99 | 99 | 0 | 33 | 19 | 0 | 0 | 0 | 0 | 0 | 0 | Nia H99 |
| 3 Peret II-26 | 6 | 99 | 99 | 99 | 0 | 2 | 0 | 12 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret III-28 | 6 | 99 | 99 | 99 | 0 | 0 | 0 | 15 | 0 | 0 | 33 | 0 | 0 | Nia H99 |
| 3 Peret III-19 | 6 | 99 | 99 | 0 | 99 | 99 | 0 | 26 | 0 | 0 | 0 | -6 | 0 | |
| 3 Peret III-19 | 6 | 99 | 99 | 0 | 99 | 33 | 0 | 9 | 19 | 0 | 0 | -3 | 0 | Car U O99 |
| 3 Peret II-26 | 6 | 99 | 99 | 0 | 99 | 0 | 0 | 24 | 19 | 0 | 0 | -3 | 0 | |
| 3 Peret III-19 | 6 | 99 | 99 | 0 | 0 | 99 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret III-19 | 6 | 99 | 99 | 0 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret IV-1 | 6 | 99 | 99 | 0 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret III-19 | 6 | 99 | 0 | 99 | 99 | 99 | 0 | 23 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret III-19 | 6 | 99 | 0 | 99 | 99 | 33 | 19 | 0 | 0 | 0 | 0 | 0 | 0 | Asc H99S>32 |
| 3 Peret III-22 | 6 | 99 | 0 | 99 | 99 | 13 | 0 | -5 | -3 | 0 | 0 | 0 | 0 | |
| 3 Peret III-19 | 6 | 99 | 0 | 99 | 20 | 33 | 19 | 0 | -6 | 0 | 0 | 0 | 0 | Asc H99S>32 |
| 3 Peret II-26 | 6 | 99 | 0 | 99 | 89 | 0 | 0 | -5 | -3 | 0 | 0 | 0 | 0 | |
| 3 Peret III-19 | 6 | 99 | 0 | 99 | 0 | 99 | 0 | 0 | 0 | 0 | 8 | -3 | 0 | Nia H99 |
| 3 Peret III-22 | 6 | 99 | 0 | 99 | 0 | 42 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret III-22 | 6 | 99 | 0 | 99 | 0 | 17 | 0 | -5 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret II-26 | 6 | 99 | 0 | 99 | 0 | 0 | 0 | -5 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret III-19 | 6 | 99 | 0 | 0 | 99 | 99 | 0 | 16 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret III-22 | 6 | 99 | 0 | 0 | 99 | 54 | 0 | -6 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret III-19 | 6 | 99 | 0 | 0 | 99 | 33 | 0 | 9 | -6 | 0 | 0 | -3 | 0 | |
| 3 Peret III-22 | 6 | 99 | 0 | 0 | 99 | 19 | 0 | -5 | -3 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret IV-1 | 6 | 99 | 0 | 0 | 99 | 0 | 0 | 14 | -3 | 0 | 0 | -3 | 0 | |
| 3 Peret III-19 | 6 | 99 | 0 | 0 | 0 | 99 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret III-22 | 6 | 99 | 0 | 0 | 0 | 63 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret III-19 | 6 | 99 | 0 | 0 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret III-22 | 6 | 99 | 0 | 0 | 0 | 22 | 0 | 0 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret IV-1 | 6 | 99 | 0 | 0 | 0 | 0 | 0 | -5 | 0 | 0 | 0 | 13 | 0 | Pry H=00 |
| 3 Peret II-23 | 6 | 0 | 99 | 99 | 99 | 96 | 0 | 0 | 0 | 0 | 20 | -8 | 0 | Nia H99 |
| 3 Peret III-22 | 6 | 0 | 99 | 99 | 99 | 68 | 0 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 61 | -6 | 16 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 55 | -6 | 16 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 48 | -6 | 19 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 42 | -6 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret II-23 | 6 | 0 | 99 | 99 | 99 | 33 | 0 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 31 | -6 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 25 | -6 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret III-22 | 6 | 0 | 99 | 99 | 99 | 23 | 0 | 0 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 19 | 0 | 2 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 19 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 17 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 15 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 13 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 12 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 9 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 7 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 5 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 3 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 2 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 1 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret IV-1 | 7 | 0 | 79 | 89 | 89 | 61 | -6 | -19 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 99 | 0 | 0 | 15 | 19 | 0 | -13 | 0 | 0 | Car U O99 |
| 3 Peret II-26 | 6 | 0 | 99 | 99 | 99 | 0 | 0 | 15 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret II-23 | 6 | 0 | 99 | 99 | 0 | 99 | 0 | 0 | 0 | -2 | 10 | -5 | 0 | Nia H99 |
| 3 Peret III-22 | 6 | 0 | 99 | 99 | 0 | 73 | 25 | 0 | 0 | -2 | 0 | 0 | 0 | Asc H99S>32 |
| 3 Peret II-23 | 8 | 0 | 99 | 99 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret III-22 | 8 | 0 | 99 | 99 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret IV-1 | 6 | 0 | 99 | 99 | 0 | 0 | 0 | 15 | 0 | 0 | -13 | 0 | 0 | |
| 3 Peret II-26 | 6 | 0 | 99 | 99 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret III-22 | 6 | 0 | 99 | 0 | 99 | 99 | 0 | 0 | 19 | 0 | 0 | 33 | 0 | Car U O99 Pry H=00 |
| 3 Peret II-23 | 6 | 0 | 99 | 0 | 99 | 99 | 0 | 0 | 0 | 0 | 0 | 53 | 0 | Pry H=00 |
| 3 Peret II-23 | 6 | 0 | 99 | 0 | 99 | 33 | 0 | 0 | 19 | 0 | 0 | -3 | 0 | Car U O99 |
| 3 Peret III-22 | 6 | 0 | 99 | 0 | 99 | 32 | 0 | 0 | 19 | 0 | 0 | -3 | 0 | Car U O99 |
| 3 Peret IV-1 | 6 | 0 | 99 | 0 | 99 | 0 | 0 | 18 | 19 | 0 | 0 | 0 | 0 | Car U O99 |
| 3 Peret II-23 | 6 | 0 | 99 | 0 | 0 | 99 | 0 | 30 | 0 | -2 | 0 | -13 | 0 | Bio H=00 O=00 S>94 |
| 3 Peret III-22 | 6 | 0 | 99 | 0 | 0 | 95 | 0 | 60 | 0 | 60 | 0 | -3 | 30 | Fol O=00 S>84 Thi H=00 O=00 S>94 |
| 3 Peret II-23 | 6 | 0 | 99 | 0 | 0 | 33 | 0 | 0 | 0 | -2 | 0 | -3 | 0 | |
| 3 Peret III-22 | 6 | 0 | 99 | 0 | 0 | 31 | 0 | 0 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret IV-1 | 6 | 0 | 99 | 0 | 0 | 0 | 0 | 15 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 99 | 99 | 99 | 0 | 0 | 0 | 0 | -5 | 0 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 99 | 99 | 89 | 0 | 0 | -6 | 0 | -2 | 0 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 99 | 99 | 33 | 0 | 0 | -6 | 0 | 0 | 0 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 99 | 99 | 28 | 0 | 0 | -6 | 0 | 0 | 0 | 0 | |
| 3 Peret II-26 | 6 | 0 | 0 | 99 | 99 | 0 | 0 | -5 | -3 | 0 | 0 | 0 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 99 | 0 | 99 | 0 | 0 | 0 | 0 | -5 | 0 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 99 | 0 | 84 | 30 | 0 | 0 | 50 | 20 | 0 | -20 | Asc H99S>32 Fol O00S>84 Nia H99 |
| 3 Peret II-23 | 6 | 0 | 0 | 99 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 99 | 0 | 27 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret II-26 | 6 | 0 | 0 | 99 | 0 | 0 | 0 | -5 | 0 | 0 | 0 | 0 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 0 | 99 | 99 | 0 | 0 | 0 | 0 | 0 | -14 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 0 | 99 | 79 | 0 | 0 | -6 | 0 | 0 | -8 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 0 | 99 | 33 | 0 | 0 | -6 | 0 | 0 | -3 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 0 | 99 | 25 | 0 | 0 | -6 | 0 | 0 | -3 | 0 | |
| 3 Peret II-26 | 6 | 0 | 0 | 0 | 99 | 0 | 0 | -8 | -3 | 0 | 0 | 0 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 0 | 0 | 99 | 0 | 0 | 0 | 0 | 0 | -13 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 0 | 0 | 72 | 0 | -20 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret II-23 | 6 | 0 | 0 | 0 | 0 | 33 | 0 | 0 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret III-22 | 6 | 0 | 0 | 0 | 0 | 23 | 0 | 0 | 0 | 0 | 0 | -3 | 0 | |
| 3 Peret II-26 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | -5 | 0 | 0 | 0 | -3 | 0 |
Vitamin Tests
A rigourous, but boring, approach would be to hold all but one variable constant, and then increment the target variable both up and down, and record what happened. Fortunately it's not possible to peg any of the intermediate values.- Ingredients are added in lots of 20 units
- Dung and Urine decrement by 10, 20, or 30 units depending on conditions
- H2O and Oxygen always decrement by 10 units
- Sunshine does its own thing
- Some conversions don't require either Dung or Urine
- Some conversions transfer between vitamins
- Some conversions are in equilibrium, with the same amount coming in and going out, and the associated vitamins may appear to be static, while in truth they're right in the thick of the fight.
- Conversion ratios vary from 3:1 to 1:5
- Mass is not conserved so far as we can tell
So ....
It's more fun anyway to chase how to force a particular vitamin to increase or decrease, and then watch for the shift points when things happen, and perhaps revisit the test later on with an alternate strategy. The data in the next table is taken from the same worksheet as the above data, and it's indexed the same way, but now we're showing the Rate of Change for each vitamin, and only including dynamic states or, in some cases, the adjacent states to show a boundary condition.
We've also added an artifical E = Environmental input, to account for self-starting conversions, thereby allowing H2O, Oxygen, and Sunshine to be treated as conditionals rather than also showing them as inputs.
You need to understand how we're using the following data, in order to understand the organization of the table. We're using the Excel Autofilter capability to isolate all conditions that can cause a particular Vitamin to increase or decrease, and then we're trying to discern the (one or more sets of) ambient conditions that are required for that particular conversion.
Some of the conversion are reversible, while other conversions create loops in the state diagram. This series of tests focuses on recipe driven changes. These testa general require an initial setup period to convert the directly controllable ingredients into vitamin levels that are sufficient to support a mid-recipe change in strategy, plus enough additional steps to delineate the behavior of any state loop.
The objective in this portion of the research is to isolate a set of 20 or so rules that are common to all states, and to plug those ambient requirement rules into the state diagram, thereby creating a state diagram for Tale 3.
| Step | Nit | Pot | H2O | Oxy | Sun | Asc | Bio | Car | Fol | Nia | Pry | Thi | Comment |
Calibration Recipes
Only about 80% of the available conversions are active on any given day (weather matters), and even the active conversions depend on the amount of sunlight that is available. The following recipes will allow you to reliably determine which of the possible conversions are currently available.
| Step | Nit | Pot | H2O | Oxy | Sun | Asc | Bio | Car | Fol | Nia | Pry | Thi | Comment |
Modern Sheep Pen Recipes
| Step | Nit | Pot | H2O | Oxy | Sun | Asc | Bio | Car | Fol | Nia | Pry | Thi | Comment |
Silkworm Recipes
| Step | Nit | Pot | H2O | Oxy | Sun | Asc | Bio | Car | Fol | Nia | Pry | Thi | Comment |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}